Article,2023-12-08,Molecular Ecology, [IF 4.9]
DOI:https://doi.org/10.1111/mec.17235
原文链接:https://onlinelibrary.wiley.com/doi/abs/10.1111/mec.17235
第一作者:Zhiyuan Ma (马志远), Shuo Jiao (焦硕)
通讯作者:Yuting Liang (梁玉婷)
合作作者:Kaikai Zheng (郑开凯), Haowei Ni (倪浩为), Dong Li (李东), Na Zhang (张娜), Yunfeng Yang (杨云锋), Jizhong Zhou (周集中), Bo Sun(孙波), Yuting Liang (梁玉婷)
主要单位:
1中国科学院南京土壤研究所土壤与农业可持续发展国家重点实验室(State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing, China)
2西北农林科技大学生命科学学院干旱作物胁迫生物学国家重点实验室(State Key Laboratory of Crop Stress Biology in Arid Areas, College of Life Sciences, Northwest A&F University, Yangling, China)
3中国科学院大学(University of the Chinese Academy of Sciences, Beijing, China)
4清华大学环境学院环境模拟与污染控制国家重点联合实验室(State Key Joint Laboratory of Environment Simulation and Pollution Control, School of Environment, Tsinghua University, Beijing, China)
5俄克拉何马大学环境基因组学研究所,土木工程与环境科学学院微生物与植物生物系(Department of Microbiology and Plant Biology, School of Civil Engineering and Environmental Sciences, Institute for Environmental Genomics, University of Oklahoma, Norman, Oklahoma, USA)
6劳伦斯伯克利国家实验室,地球与环境科学(Earth and Environmental Sciences, Lawrence Berkeley National Laboratory, B
- 摘要 -
研究不同空间尺度下土壤微生物功能异质性及其与土壤碳矿化的关系,是预测土壤碳稳定性对环境变化和人为干扰响应的重要手段。利用MiSeq测序和GeoChip基因芯片技术,对中国典型水田土壤样品429份进行了分析,分析了土壤中细菌和真菌群落以及与碳矿化相关的功能基因。我们假设细菌和真菌碳矿化产生的二氧化碳排放取决于它们各自的碳消耗策略,而碳消耗策略依赖于生物多样性和功能基因之间相互作用的调节。结果表明,从数百米到数千公里的空间尺度,真菌群落的空间周转是细菌群落的2-4倍。空间尺度对微生物群落组成的影响大于其对微生物群落功能特征的影响。此外,基于不同空间尺度的功能网络构建,我们发现与碳矿化相关的前10个分类群中的细菌和真菌在区域尺度上都表现出通才物种的普遍存在。该研究强调了空间尺度格局在土壤细菌和真菌碳降解功能中的重要性,加深了我们对微生物分解者与土壤异质性之间的关系如何影响碳矿化和土壤碳矿化的认识。
- 引言 -
土壤微生物是分解和参与陆地碳源汇动态的重要引擎。大多数土壤微生物是异养的,它们以消耗土壤有机碳(SOC)为能量,这种碳矿化过程可以加速土壤向大气排放CO2。土壤碳矿化是碳循环的关键过程,与土壤碳质量直接相关。土壤微生物群落的特殊特征,包括结构和功能特征,会导致碳分解的变化。细菌和真菌是分解群落的两个主要群体,它们表现出多种影响土壤碳固存和陆地碳稳定性的策略。例如,细菌是简单碳水化合物、有机酸和氨基酸的主要分解者,而真菌在分解难降解土壤碳方面比细菌更重要。对这些微生物所采用的不同碳分解策略的细化研究可以更全面地理解微生物介导下的土壤-气候反馈是如何运作的。然而,异质性对微生物功能生态学的许多方面的潜在深远影响,尚未被很好地理解或纳入我们对生态系统在多空间尺度上响应气候变化的功能的认识。
生态学中的空间尺度表示邻近的相互有效影响的生物地理范围。物种共存对生态系统功能至关重要,它依赖于空间尺度,这种关系与跨景观的群落多样性管理有关。生态模式的尺度依赖性通常是由抽样效应引起的:增加观测尺度可以增加资源的存在和利用数量,从而使物种与观测尺度呈单调正相关。Bruelheide et al (2018)发现,群落功能特征的组成和驱动力在局地区域(1-1000 m2)和大面积区域(25 - 12500 km2)之间存在差异,表明生态模式依赖于空间尺度,呈现出特征驱动情景。然而,不同微生物类群的空间尺度及其功能特征对维持生态系统生物地球化学循环是否具有重要意义尚不清楚。土壤中碳降解微生物呈分形,在一定尺度范围内表现出自相似性。阐明群落相似性在空间尺度上的差异的重要性、量化不同群落组成对碳矿化过程的影响,应该与生态功能联系起来。
距离衰减关系(distance decay relationship, DDR)是生态学中一种常见的生物地理模型,指的是群落相似性随地理距离的减小。已有学者对不同区域范围的微生物群落组成的DDR进行了调查,包括大陆尺度和全球尺度两个方面。这种关系在不同的微生物分类群中有所不同。通过对1 m-700 km范围内重氮营养型土壤和细菌群落的DDR进行研究,发现重氮营养型土壤的DDR斜率(空间周转率)大于细菌。导致不同DDR模式的微生物群落组成的影响因素,一般认为是选择和分散以及漂移和多样化起着重要的作用。在pH、SOC和NH4+含量等因素的驱动下,环境过滤会影响土壤微生物的空间模式。概率分散可以独立于生态位差异影响DDR模式。目前,土壤微生物的研究正从群落组成向功能性状发展。然而,不同微生物群的空间尺度和功能特征对土壤碳稳定性的影响尚不清楚。
全球水稻土面积达1.5亿公顷,是世界上最重要的农业生态系统之一。这一广阔的分布区域为在大空间尺度上研究微生物功能性状提供了理想的栖息地。在这项研究中,我们从中国水稻种植区的稻田中收集了429个土壤样本。通过16S rRNA和ITS扩增子测序以及高通量功能基因阵列GeoChip分析土壤细菌和真菌群落以及碳降解功能基因。采样采用了空间设计,实现局地尺度(1 ~ 100 m)、中尺度(0.5 ~ 50 km)、区域尺度(100 ~ 3500 km)和整体尺度(1 ~ 3500 km)对距离的均衡分布。在此,我们提出不同微生物对碳矿化的参与取决于细菌和真菌在不同空间尺度上表现出的碳消耗策略,这些策略受到生物多样性和功能基因之间权衡的影响。我们的研究表明,土壤细菌和真菌碳降解功能的空间尺度模式的重要性,可以加深对微生物分解者和土壤异质性之间的关系如何影响碳矿化和随后的温室气体排放的理解。
- 结果 -
土壤碳矿化与细菌和真菌群落结构及碳降解功能基因的分布
Soil carbon mineralization and the distribution of the bacterial and fungal community structure and carbon degradation functional genes
对中国稻田的所有429个样品进行了土壤碳矿化测定(图1a)。这些不同的碳矿化模式与地理格局、气候条件、农业实践、土壤类型和微生物活动等复杂的环境特征密切相关。东北3个样点(HL、CC、SY)土壤碳矿化程度分别为16.31±2.71、42.77±27.35、23.92±17.53 mg/(kg d);华北平原(YY和FQ)分别为58.66±24.31和45.29±8.81 mg/ (kg d);长江中下游(HY、ZX、QZ、LA)呈波动趋势(分别为40.57±23.82、16.27±8.57、57.76±17.49、29.39±17.04 mg/ (kg d));华南地区(CT、JO、HK和QX)从38.07±20.61依次增加到40.57±23.82、41.84±16.45和52.14±24.18 mg/(kg d)。以土壤属性为例,土壤pH值为3.56 ~ 8.65,DOC值为57.15 ~ 509.83 mg/kg。代表土壤异质性的土壤地球化学变量随空间距离的增加而增加,即局地尺度<中尺度<区域尺度。使用Pearson相关分析C矿化与土壤性质和微生物生物量之间的关系(图1b)。在各地球化学属性中,碳矿化与土壤pH和DOC呈显著正相关(P< 0.05),与细菌生物量(P= 0.001)和真菌生物量(P= 0.033)呈高度相关。
在所有429个样本中,细菌和真菌的总测序读取数分别为24,291,099和12,461,277。为了最大限度地减少不同样品的读取计数变化的影响,对细菌和真菌的所有样品分别抽平到每个样品的20,000和10,000个读取。共鉴定出细菌群落的70,145个操作分类单元(OTU, 97%相似度)和真菌群落的49,165个OTU(97%相似度)。土壤微生物群落多样性在4个空间尺度上呈现异质性。其中,Proteobacteria (30.28±4.66%)为优势菌门,Cyanobacteria (14.30±10.25%)次之。Mortierellomycota (54.67±26.30%)为优势真菌门,其次为Ascomycota (19.58±11.20%)。土壤微生物群落表现出明显的区域分布特征(图2a)。土壤细菌和真菌的组成均表现出明显的区域差异,与气候带相关。
为了更好地了解微生物功能基因如何影响不同气候区有机碳的分解,我们对参与各种碳循环过程的关键功能基因进行了GeoChip 5.0阵列高通量筛选(图2b)。对所有水稻样品的碳降解功能基因进行了分析。结果表明,细菌(19个核心基因)和真菌(19个核心基因)参与碳降解的功能基因丰度存在显著差异。例如,细菌组的功能基因更多地归因于编码淀粉降解基因的蛋白质,包括apu (8.77±0.36,相对基因丰度)、cda (568.04±1.17)、nplT (500.24±3.90)和pula (764.29±1.45),而真菌组的功能基因更多地与纤维素、半纤维素、几丁质和木质素等难降解碳化合物的降解相关,包括exoglucanase (554.59±1.11)、xyla (1.52±0.22)。deacetylase (737.79±1.19),ligninase (1074.98±2.00),mnp (2210.32±3.61)。此外,我们使用Pearson’s correlation分析了细菌和真菌多样性与其各自碳功能基因之间的关系。
细菌的β多样性与碳分解基因呈显著正相关(14/19),α多样性与功能基因无显著差异。真菌碳分解功能基因与β多样性呈显著正相关(11/19),与α多样性呈显著负相关(2/19)。研究结果表明,细菌和真菌与碳功能基因(C功能基因)丰度在空间尺度上呈显著正相关,而不是在单个采样点内。此外,与碳分解相关的大部分功能基因,如amyA、glucoamylase、ara和glx,在4个气候区表现出显著差异,细菌和真菌的反应也不完全一致。结果表明,微生物群落的一系列特定性状和功能可能与不同的碳代谢途径有关。
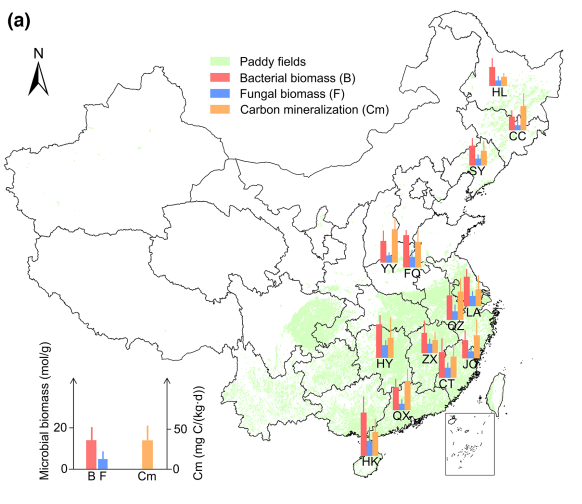

图1 中国典型稻田碳矿化情景及非生物因子与生物因子的显著相关性
(a)不同纬度的碳矿化和细菌、真菌生物量。柱状图为13个采样点33个平行土壤样品(3个重复,11个生物重复)的细菌生物量(mol/g)、真菌生物量(mol/g)和碳矿化(mg/(kg d))的平均值,误差条为标准差;
(b)基于Pearson相关分析的碳矿化与pH、溶解有机碳(DOC)、细菌生物量和真菌生物量的关系。阴影区域表示95%置信区间
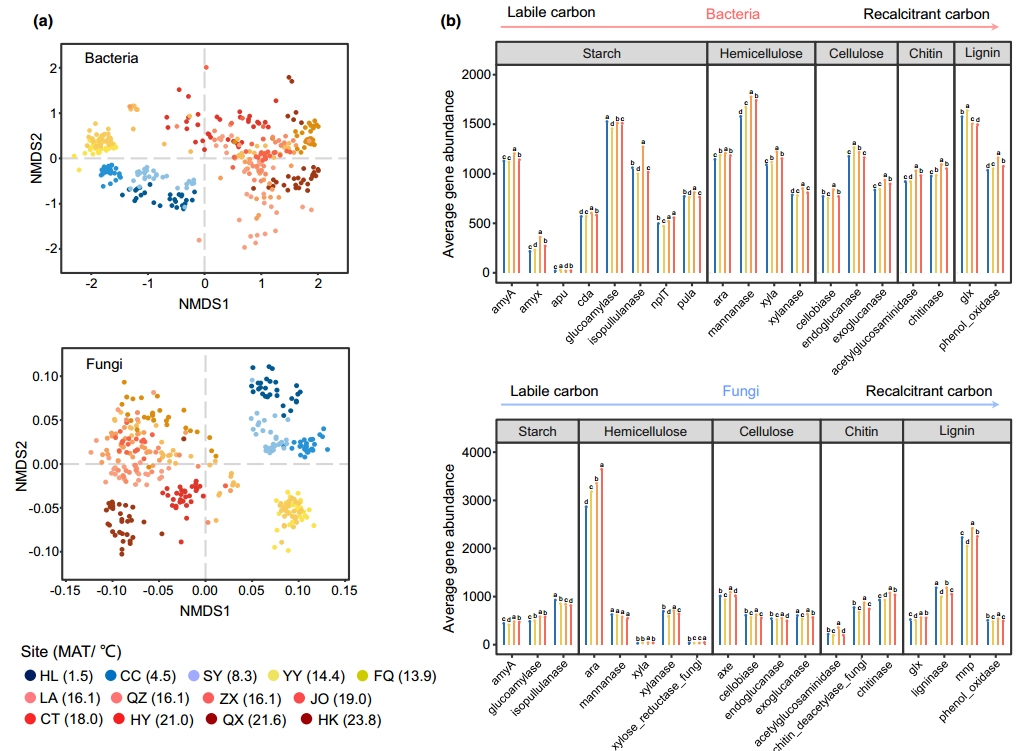
图2 细菌和真菌群落结构与碳矿化相关的功能基因丰度。
(a)基于布雷-柯蒂斯距离的细菌(上图)和真菌(下图)群落的非度量多维尺度(NMDS)。从深蓝色(HL)到红色(HK)的颜色代表了从47.579°N到19.758°N的中国北部到南部的样本。
(b)列表示在中温带(蓝色)、暖温带(黄色)、亚热带(橙色)和热带(红色)温度下,细菌(上)和真菌(下)中参与淀粉、半纤维素、纤维素、几丁质和木质素分解的功能基因的平均丰度。误差条表示标准差。误差条上方显示了基于单因素方差分析(多重比较,Fisher’s LSD test)的各气候带配对组基因丰度的显著差异(P< 0.05)
从局地到区域尺度土壤微生物群落结构和功能基因的空间迁移
Spatial turnovers of the soil microbial community structure and functional genes from local to regional scales
在局地尺度1 ~ 100 m、中尺度1 ~ 50 km、区域尺度100 ~ 3500 km和全局1 ~ 3500 km四个空间尺度上,研究了土壤微生物群落的距离衰减关系。细菌和真菌群落的群落相似性与地理距离呈显著的线性回归关系(P< 0.05)。01、图3a、b)。在所有空间尺度上,真菌群落的空间周转率都是细菌群落的2 ~ 4倍。在门水平上进一步计算不同微生物分类的空间更替。不同门间DDR周转的空间尺度依赖性与整个群落的空间尺度依赖性基本一致。利用DDR线性最小二乘回归的斜率计算空间周转率,揭示了群落异质性的层次格局,细菌和真菌群落在更大的空间尺度上表现出更高的异质性(区域尺度>全局尺度>中尺度>局地尺度)。
然后,我们分析了与碳分解相关的功能基因水平上的微生物功能异质性(图3c,d)。微生物功能基因的变异,如amyA、乙酰氨基葡萄糖酶和外葡聚糖酶,随着地理距离的增加而增加。与群落结构模式相反,细菌在区域尺度上和真菌在中尺度上的功能异质性最高。这些结果表明,参与碳分解的功能基因随着空间距离的增加而变化。采用随机森林分析方法研究了环境因子对细菌和真菌α和β多样性的影响。微生物α多样性的主要驱动因素在四个空间尺度上有所不同。土壤NH4−-N(13.4%)和TP(43.5%)分别是局地和中尺度上细菌数量的主要影响因子。土壤NO3−-N在中尺度上对真菌多样性贡献最大(24.8%),在局地尺度上对真菌多样性贡献不显著。气候因子对细菌和真菌α多样性的影响随空间尺度的增大而增大,在区域和整体尺度上,MAP和MAT对细菌和真菌α多样性的影响最大。相比之下,土壤pH值在四个空间尺度上是细菌β多样性的最重要驱动因素。在区域和整体尺度上,MAP和MAT对真菌β多样性的影响增强,贡献最大。
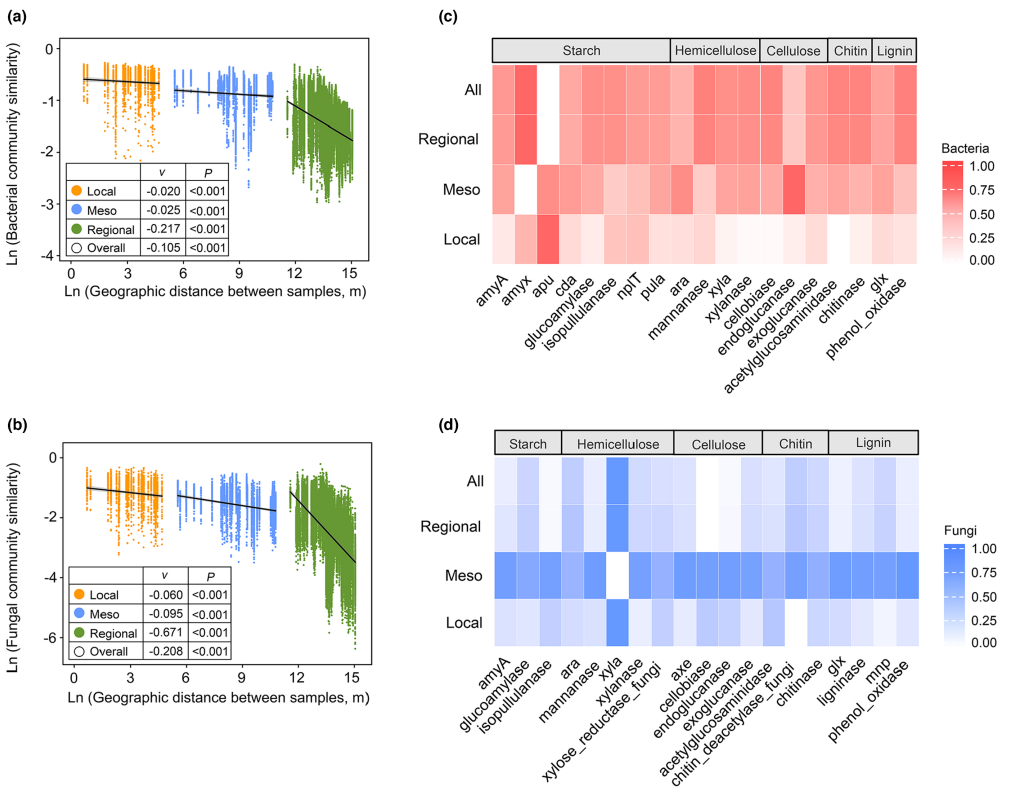
图3 四个空间尺度下土壤细菌和真菌群落结构和碳功能特征的距离衰减规律。
(a)不同空间尺度下基于Bray-Curtis距离的土壤细菌群落距离衰减关系(DDRs)散点图。斜线斜率(v)表示微生物空间周转率。局地尺度:1 - 100 m;中尺度:1-50km;区域尺度:100-3500km;全局尺度:1 m – 3500km。
(b)不同空间尺度下土壤真菌的DDR。
(c)细菌碳功能基因在不同空间尺度上的变异热图。
(d)真菌碳功能基因在不同空间尺度上的变异。
土壤微生物结构和功能特征与土壤碳矿化的联系
Linkage of soil microbial structural and functional traits to soil carbon mineralization
考虑到生物多样性和功能基因对碳矿化的贡献会受到生物特征和环境因素的影响,包括气候变量(MAT和MAP)以及环境参数(DOC和pH),我们将这些因素与α/β多样性和碳分解基因数据一起纳入我们的模型。细菌多样性(α和β多样性)在四个空间尺度上对碳矿化的影响大于真菌多样性(图4a,b)。细菌和真菌的α和β多样性在中尺度上贡献最大。促进碳矿化的重要基因群是细菌群落的amyA、葡萄糖淀粉酶和几丁质酶,真菌群落的ara、纤维素酶、几丁质脱乙酰酶、木质素酶和mnp(图4c、d)。这一结果也表明细菌功能基因主要影响易分解碳部分,而真菌类群主要影响顽难分解碳部分。然后,我们使用偏最小二乘路径分析方法(PLS-PM)构建了生物多样性(α和β多样性)、功能基因和异位呼吸(等于碳矿化)的结构方程模型(SEM)。通过SEM分析,不同尺度下细菌碳功能基因的相关系数为0.96 ~ 0.97 (P< 0.001)。中尺度上,真菌碳功能基因与碳矿化的相关系数为- 0.02 (P< 0.001)。而空间尺度变化对物种多样性的影响更为明显。在中尺度上,细菌和真菌的β多样性对碳矿化的贡献为0.25。随着尺度的增大,β多样性的贡献减小到小于0.001。这些结果表明,空间尺度对微生物群落功能特征的影响呈现出异步变化的特征,空间尺度效应对微生物群落组成的影响大于对微生物群落功能特征的影响。构建协同网络,将土壤碳矿化与土壤结构功能特征在不同空间尺度上联系在一起(图5,表1)细菌类群在综合功能网络中的优势度随空间尺度呈比例增加(从63.20%增加到83.67%)。这一观察结果与随机森林分析结果一致,表明细菌比真菌对碳矿化的贡献更大。在中尺度上,该网络的节点数最多(338),边数最多(6277),网络异质性值为0.924。然而,随着空间尺度的增大,进入网络的节点总数明显减少。明显的结果是没有碳分解功能基因在区域和整体尺度上成功整合。根据局地尺度、中尺度和区域尺度子网的加权排名选择前10个OTU。随着空间尺度的扩大,多面手微生物的丰度显著增加。值得注意的是,在区域尺度上,前10个分类群中的细菌和真菌都是通才物种。这一发现表明,随着空间尺度的增加,微生物相互作用变得更加频繁,多面手可以通过功能互补增强其碳分解能力。这一发现还强调了空间尺度对微生物群落组成的主要影响,超过了对与碳分解相关的功能基因的影响。

图4 生物多样性和功能基因多样性对碳矿化过程的贡献。
(a) α多样性和(b) β多样性对四个空间尺度碳矿化的贡献采用随机森林(RF)分析计算细菌和真菌多样性对碳矿化的平均预测值(均方误差(MSE)增加百分比)。
(c)细菌和(d)真菌中功能基因对碳矿化的贡献用平均预测值表示。各预测因子的显著性水平分别为:* P < 0.05;* * P < 0.01。

图5 四个空间尺度的碳矿化共生网络分析。
网络途径显示了功能细菌和真菌群落与碳矿化相关基因之间的基本联系。在四个空间尺度上,每个节点(红色或蓝色点)代表一个细菌或真菌的操作分类单位(OTU,在属水平上),有助于碳矿化(Cm),每个边代表一个积极或消极的相互作用。每个节点的大小表示Cm的量级,每个边缘的渐变颜色表示显著调节的权重。粗体字表示根据体重秤对Cm贡献最大的10个OTU(细菌或真菌)。种*表示在每个子网中检测到的概率超过50%的通才分类单元。斜体字代表细菌(紫色点)和真菌(绿色点)调节Cm过程的关键基因
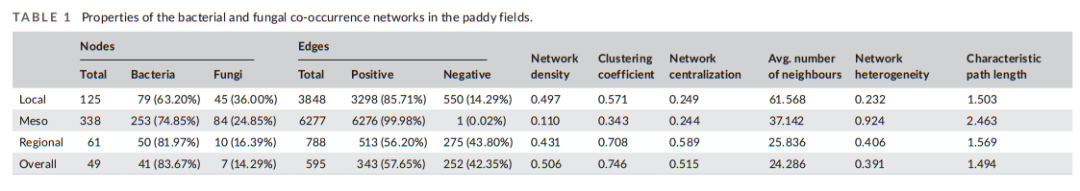
表1 水田细菌和真菌共生网络的特征
- 讨论 -
生物多样性变化与功能性状变化之间关系的强弱取决于空间尺度。在本研究中,土壤细菌和真菌在门水平上的空间周转速率与整个群落的空间尺度效应大致一致(图3 a、b;)。然而,细菌和真菌之间与碳分解相关的功能基因差异不太依赖于空间周转,细菌和真菌对碳功能基因的贡献最显著的是中尺度。细菌α多样性与碳功能基因的相关性随着养殖规模的增加而显著增加。真菌多样性(α和β)在中尺度上保持最显著的相关性。这些结果表明,空间尺度效应对微生物群落功能性状的影响是异步变化的,空间尺度效应对细菌群落组成的影响大于对其功能特征的影响。
为了解释生物多样性与碳矿化功能性状之间的非同步变化,我们提出了两种理论。
第一个是概率分散影响局部动力学。Thompson等人(2017)表明,适度的扩散可以允许物种在最优环境中有效地跟踪空间变化并增加其功能。先前的研究表明,当个体直径大于20 μm时,微生物的分散概率降低,而直径小于约1 μm的微生物不受分散。中性模型表明,在局地和中尺度上,均质分散(66.43%和69.23%)和异质性选择(32.40%和30.77%)占主导地位。在区域尺度上,影响细菌群落组成的非显性过程较多(57.69%)。这导致同质分散(17.95%)和异质选择组成(20.51%)的减少。细菌比真菌更容易受到扩散限制,因为它们的体积更小(尺寸假说),导致细菌在多个尺度上比真菌具有更高的平均功能丰度。移动消费者从一个补丁移动到另一个补丁可以稳定功能。
第二个假设是土壤异质性增加了环境条件的范围(环境过滤)。异质性是土壤的基本特性,是土壤微生物多样性出现和维持的基础。此外,非均质土壤也导致土壤有机质的化学异质性,在景观尺度上高产农田也是如此。由可变的非生物因素施加的环境过滤导致高或低的群落转换率,这取决于这些因素的一致性。在本研究中,环境因子(DOM和pH)对细菌α-多样性的影响在各个空间尺度上都大于真菌。DOM影响土壤中基本的生物地球化学过程,如养分循环和有机质储存。土壤微生物组对DOM组成有很强的影响,细菌对DOM组成的影响大于真菌。我们假设一些真菌,如外生菌根真菌,优先降解大的衍生聚合物(如木质素),部分矿化并将其转化为一系列不同的小分子,随后在微生物加工过程中被细菌消耗。这一过程对真菌的周转率要求高于细菌,导致细菌多样性增加。
土壤pH值是影响细菌和真菌群落的另一个关键因素。与细菌相反,土壤真菌的空间分布与土壤pH值有关,可能是因为真菌比细菌有更宽的pH适应范围。因此,在局地空间尺度上,细菌比真菌对环境过滤更敏感(群落周转低)。相比之下,最近的一项研究表明,在北方森林土壤中,细菌群落的空间周转大于真菌群落的空间周转。影响距离衰减关系(DDR)的潜在因素(环境变量或地理距离)的相对重要性在不同的生境(如高山草原、沙漠、荒漠草原、典型草原和整个样带)中也有所不同。森林和竹林等自然生态系统较少受到人类的干扰。因此,土壤真菌群落比土壤细菌群落更具植物特异性,尤其是根际真菌。水田是典型的人类管理生态系统,由于现代农业活动,细菌扩散限制可能会降低。
在多个空间尺度上,细菌对碳矿化的贡献大于真菌,这与细菌的功能特性一致。无论群落α或β多样性如何,随机森林结果表明,细菌参与碳代谢的程度明显大于真菌(图4a,b)。细菌和真菌在空间尺度效应诱导下的共存模式是中尺度生物评价的可靠指标。
由于不同空间尺度上微生物多样性对生境类型的潜在依赖性,微生物往往表现出对局部资源区的特征性专业化分解,或对更广泛的化合物的通用性分解。我们进一步分析了细菌和真菌代谢不同碳源的潜力,从易分解碳到难分解碳。我们发现细菌群落对易分解碳 (amyA和葡萄糖淀粉酶)贡献更多,而真菌则更多地参与难分解碳的降解(如,ara,纤维素酶,几丁质脱乙酰酶,木质素酶和mnp)(图4c,d)。即使对难分解碳的代谢,细菌对相关功能基因的贡献也是真菌的1.2倍。这一发现与之前的研究结果一致,细菌是简单碳水化合物、有机酸和氨基酸的主要分解者,而真菌对难降解土壤碳的分解更为重要。与真菌相比,细菌更大的系统发育多样性和代谢能力的广度似乎对碳的分解有更大的影响。
细菌和真菌具有不同的底物偏好和代谢差异,真菌或细菌共同发挥不同的作用是理解土壤碳矿化的核心。在本研究中,中国稻田中细菌和真菌表现出多种潜在的微生物相互作用。在四个空间尺度上,与碳代谢相关的高丰度细菌和真菌的比例分别为:局地尺度(细菌= 63.20%,真菌= 36.00%)和中尺度(细菌= 74.85%,真菌= 24.85%)、区域尺度(细菌= 81.97%,真菌= 16.39%)和整体尺度(细菌= 83.67%,真菌= 14.29%)(图5和表1)。在共生网络中,细菌与碳矿化的关联比真菌更强,这与细菌在多尺度上的功能性状强度高于真菌是一致的。
细菌和真菌共享相同的栖息地,因此几乎可以肯定在土壤中经常相互作用。因此,基于共存理论,更大的空间尺度涵盖了更大范围的环境,这增加了物种进行生态位划分的机会,并促进了空间生态位的互补性。我们推测,在这些空间尺度的碳过程中,正或负共现关系可能代表了个体之间互补或竞争的潜在相互作用。竞争作为主要结构性力量的重要性可能显现;同时,群落可能由几个特异微生物组成,每个特异微生物消耗有限范围的底物,从而减少了群落中每个成员之间的竞争,增加了互补性。当按权重排序前10名细菌或真菌时,随着空间尺度的增加,特异微生物分类群的数量明显下降。Gravel等人(2011)发现,广谱微生物组合在一系列碳基质上的生产力更高,因为它们更有能力利用资源环境中施加的异质性,而特异微生物组合的生物多样性-生态系统功能(BEF)关系的斜率更强,因为生态位互补性增强。细菌和真菌群落可以通过土壤生态位内部和土壤生态位之间的互补来支持碳代谢,这意味着空间尺度效应可以对碳矿化产生积极影响。
土壤有机碳(SOC)是一种不可再生资源,目前其消耗速度快于形成速度。土壤条件或管理实践的任何变化都会改变地球化学或环境化学过程,从而影响农业生态系统中碳和氮的循环,最终导致温室气体(即CH4、N2O和CO2)的产生。在过去的50年里,由于发展和采用改良品种和管理以及技术进步,作物特定产量增加(全球干生物量产量增加240%),北半球的大气二氧化碳增加了50%。越来越多的证据指出土壤微生物的作用,土壤微生物是分解和参与陆地碳源汇动态的重要引擎。在本研究中,pH和DOC是与碳矿化呈正相关的两个非生物因子。两者都通过一系列氧化还原反应参与微生物介导的碳转化过程,交换或提供游离电子产生新的氧化剂(如O2、NO3−、Mn4+、Fe3+和SO42−),并因此排放温室气体。然而,在大规模的研究中,我们通常忽略了微生物对温室气体排放的贡献。这是因为虽然这些研究可以在局地尺度上测试潜在的机制,但它们也不能直接解决更广泛尺度上的理论预测。多尺度测量可用于预测与观测数据固有功能相关的微生物多样性转换的空间模式,特别是在大空间尺度上。土壤异质性和空间尺度效应对微生物群落组成和功能性状具有显著的非同步效应。它们还可以在基于生态位互补性的大空间尺度上驱动同步模式。利用更大的数据集,包括未来跨越空间位置网络的时间序列,我们可以增加对与多样性和碳矿化功能共变的非控制变量的信心,并表征不同水平上物种波动的同步和交叉相干尺度。
我们的研究比较了从局地尺度到全局尺度四个空间尺度上细菌和真菌的生物地理格局和碳分解趋势。pH和DOC是与碳矿化呈正相关的两个非生物因子。微生物性状的权衡决定了微生物有机碳投入生物合成的比例。基于DDR,细菌在不同空间尺度上的扩散能力大于真菌,这可以合理解释微生物群落与土壤有机质代谢之间的关系。细菌群落对碳矿化的贡献更大,因为细菌的分解代谢广度更集中于易分解碳,而真菌则更多地参与难分解碳的降解。功能性状和微生物群落受空间尺度效应的影响,表现出显著的非同步性。生态位互补可以使功能性状均一化,促进微生物对碳的代谢。由于土壤资源有限,该群落由几个特异微生物组成,每个专家消耗有限范围的基质。然而,随着空间尺度的增加,不同的多面手群体之间的相互作用不断升级,导致碳分解过程加速。总之,我们的研究为预测空间群落生态中最多样化和最复杂的微生物的独特功能特征提供了新的见解。
参考文献
Ma, Z., Jiao, S., Zheng, K., Ni, H., Li, D., Zhang, N., Yang, Y., Zhou, J., Sun, B., & Liang, Y. (2023). Multiple spatial scales of bacterial and fungal structural and functional traits affect carbon mineralization. Molecular Ecology, 00, 1–16