高等开花植物70%左右是多倍体,多倍体在我们日常生活中非常普遍,人类的主要食物面包就是异源多倍体小麦的加工产品,我们经常吃的无籽西瓜、香蕉、草莓等也都是多倍体。
多倍化是开花植物共同的进化模式,研究表明,大部分植物经历过多倍化的过程,新形成的多倍体面临的最大挑战是减数分裂过程中染色体的配对与分离,因此,多倍体的稳定必须经过一个细胞学二倍化的过程,二倍体化开始于最早的染色体组融合和加倍。作为小麦的野生近缘种,长穗偃麦草因可与小麦进行杂交在小麦遗传改良中发挥了重要的作用。长穗偃麦草按倍性可划分为二倍体,四倍体和十倍体,二倍体长穗偃麦草的染色体组命名为E组,而四倍体长穗偃麦草和十倍体长穗偃麦草的基因组组成和起源仍存在争议。
2022年12月,中国科学院遗传与发育生物学研究所韩方普研究组在《The Plant Journal》杂志上发表了题为“Autoploid origin and rapid diploidization of the tetraploid Thinopyrum elongatum revealed by genome differentiation and chromosome pairing in meiosis”(DOI: 10.1111/TPJ.16066)的研究论文,研究发现四倍体长穗偃麦草是同源起源的,在其形成后经历了快速的细胞学二倍化,但其染色体组间的分化是缓慢的。
本研究利用染色体微切割技术对长穗偃麦草的7EL染色体进行了分离,并利用DOP-PCR和斑点杂交技术获得了长穗偃麦草特异的重复序列。利用发现的特异重复序列进行荧光原位杂交可将天然四倍体长穗偃麦草分成两个染色体组E1和E2,类似于异源四倍体的两个染色体组。然而,利用具有E1和E2染色体的部分双二倍体的杂交后代进行减数分裂研究发现,在四倍体小麦背景下,在减数第一次分裂中期来源于E1和E2的染色体能够很好地配对,由于四倍体小麦中含有能够抑制部分同源染色体配对的Ph1基因,这表明E1和E2两个染色体组是同源而不是部分同源的,这一结果证明天然四倍体长穗偃麦草是同源四倍体。
为进一步探究四倍体长穗偃麦草的进化特征,我们利用秋水仙素进行加倍获得了人工合成的同源四倍体长穗偃麦草。通过减数分裂研究表明,天然四倍体和人工加倍四倍体均具有类似于二倍体的减数分裂行为,在S1代就能形成14个稳定的二价体。不同的是,与天然四倍体相比,人工合成的长穗偃麦草从S1代到S4代均没有发现染色体的分组现象。这些结果表明,同源四倍体长穗偃麦草形成后经历了快速的细胞学二倍体化,但染色体组间的分化是缓慢的。后续的研究表明无论是天然四倍体还是人工合成的四倍体长穗偃麦草,他们染色体的配对均起始于基数为7且成簇的着丝粒配对,而且优先于端粒花束的形成(文章正在整理中)。
遗传发育所韩方普组石庆华博士和郭宪瑞博士为该文章的共同第一作者,韩方普研究员和张晶博士为共同通讯作者。本项研究中染色体微切割工作与本所胡赞民实验室合作完成。该研究得到国家自然科学基金重大项目的资助。
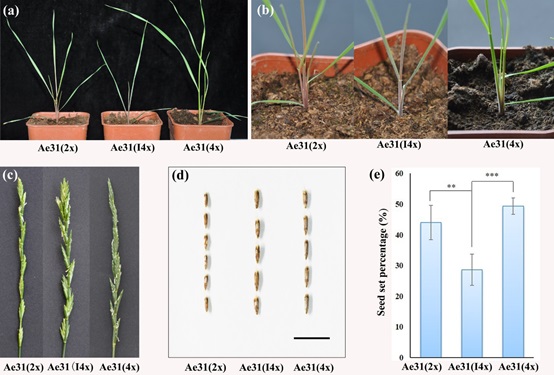
图1 :二倍体、人工加倍四倍体和天然四倍体长穗偃麦草的形态特点
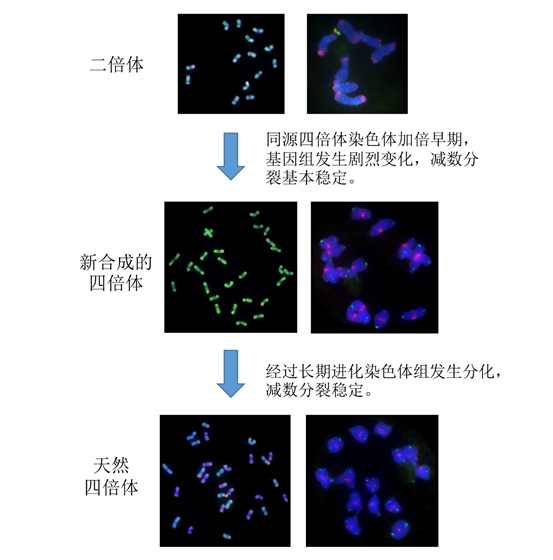
图2 :四倍体长穗偃麦草进化模式图