兜兰属(Paphiopedilum)是兰科植物最重要的属之一,其唇瓣特化成兜状或拖鞋状,故又称为“拖鞋兰”、“仙履兰”等。兜兰属植物以其奇特的花型,丰富绚丽的花色和持久的花期而具有极高的观赏价值。尽管我国兜兰属植物资源非常丰富,但大部分种类由于其自身在野生生长环境下繁殖困难,加上过度采挖以及生长环境的破坏,已经濒临灭绝。现存所有野生种均被列入《濒危野生动植物种国际贸易公约》附录I 中而被禁止交易。兜兰种子十分细小,且胚发育不完全,没有子叶和胚乳(见图1),在自然环境中需要与真菌共生才能部分萌发。在人工无菌播种中虽然提供了必要的矿质元素和其它营养物质,绝大部分兜兰的成熟种子也难以萌发。科研人员在不同胚龄种子的无菌播种实验中发现,发育前期的种子萌发率虽然相对较高,但是由于营养物质积累不充分,胚冲破种皮后,发育进程缓慢,成苗率低;而成熟后的种子营养物质积累相对充分,能满足种子萌发后初期的物质与能量需求,萌发后的成苗率较高,但种子萌发率却极低。
是什么因子影响不同成熟度兜兰种子的萌发呢?中科院华南植物园农资中心房林副研究员和硕士生徐新等科研人员,首先对包括脱落酸在内的几种内源激素的功能进行了研究。结果显示:外源脱落酸处理仅能够抑制未成熟种子的萌发,而脱落酸抑制剂处理可以降低成熟种子内源脱落酸含量,但对萌发率无显著影响。而木质素的积累与兜兰种子萌发率的下降密切相关。
木质素是由芳香类化合物单体经过脱氢聚合反应形成的复杂网状结构。从分子结构上看,木质素可以分为两大类:甲基化木质素以及非甲基化木质素,其中,甲基化木质素由愈创木基木质素 (G型木质素), 紫丁香基木质素(S型木质素)和5-羟基愈创木基木质素(5H型木质素)三种组成,非甲基化由羟基苯基木质素 (H型木质素)和咖啡基木质素(C型木质素)两种组成。甲基化的G型和S型木质素广泛的分布在植物根茎等木质化的组织中,而非甲基化木质素仅在一些兰科和仙人掌科植物的种子中。科研人员通过二维核磁、GC硫代酸解和红外光谱三种技术来分析兜兰种胚中木质素的单体结构,结果显示不同于茎干等组织,兜兰种子中富含非甲基化木质素,而富含这种木质素的种皮具有强不透水性,从而阻止水份和营养物质进入种子内部从而阻止种子萌发。
为了进一步探索兜兰种皮中非甲基化木质素的合成通路,科研人员对不同发育程度下杏黄兜兰的种子进行了转录组分析。KEGG富集分析表明,种子成熟过程中,苯丙烷生物合成和苯丙氨酸代谢相关的大量基因被差异表达。PAL,4CL,HCT和CSE上调表达与C型和H型木质素积累有关。 CCoAOMT,F5H和COMT的表达维持在低水平或下调以抑制G型和S型木质素的积累(见图2)。
非甲基化木质素在兜兰种子中积累的独特生物学意义是什么呢?在野生状态下,兜兰种子需要与适宜的真菌共生才能萌发。而非甲基化木质素本身的结构是极其稳定的,在自然界中几乎只能被真菌降解。由此,推测高度木质化的种皮是兜兰在野外状态下适应真菌共生的策略之一。
相关研究结果已发表在BMC genomics(《BMC基因组学》)(2020, 21:524)上。论文链接:https://bmcgenomics.biomedcentral.com/articles/10.1186/s12864-020-06931-1
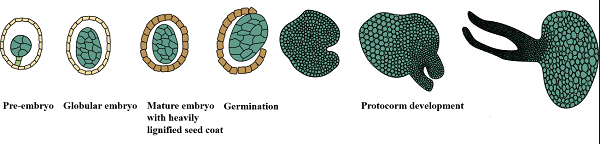
图1. 兜兰种胚发育与萌发
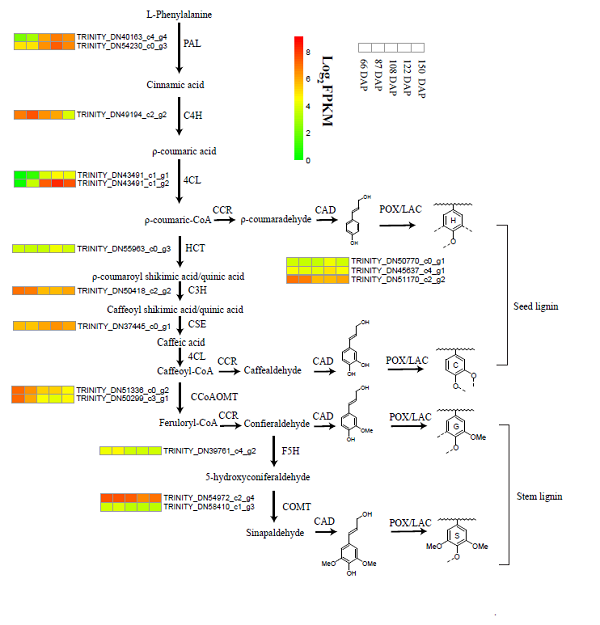
图2. 兜兰种子木质素和茎叶木质素的合成途径