太阳光不仅是植物光合作用的能量来源,也是一种重要的环境信号,调节植物的生长发育进程。其中幼苗光形态建成受光质、光量的精确调控,涉及不同的光受体和一系列信号调控因子。COP1是光形态建成的一个明星蛋白,它作为一种E3泛素连接酶,与目标蛋白互作并促进其降解,COP1在植物和动物中广泛存在。真核生物的基因大多含有内含子,转录形成前体mRNA,通过动态剪接组装,形成成熟的mRNA。可变剪接也称选择性剪接,是一种非常重要的基因转录后调控机制,在特定位点发生剪接从而生成多种mRNA,翻译成不同蛋白质,在相应的发育时期发挥重要功能。近年来,有研究通过转录组分析了光照对于可变剪接的调控现象,但对其中的具体调控机制并不清楚。
中科院植物所林荣呈研究组鉴定到RNA解旋酶UAP56是调控光形态建成的一个新因子。UAP56在拟南芥中有两个高度同源的成员UAP56a和UAP56b,位于染色体上相邻的位置。研究人员从遗传学入手,发现UAP56a和UAP56b功能冗余,但两者纯合致死,利用uap56a纯合与uap56b杂合的突变材料,证明UAP56参与到光敏色素和隐花色素光受体介导的光形态建成信号通路中,是一个负向调控因子。通过寻找互作蛋白,发现UAP56与COP1以及剪接辅助因子U2AF65直接相互作用。研究人员进一步利用Nanopore三代转录组测序技术,发现在黑暗及光照条件下,UAP56和COP1调控了大量基因的转录与可变剪接,其中包括很多植物生长发育、光合作用以及光信号因子等相关基因都受二者转录或剪接的调控。进一步进行RNA免疫沉淀实验,显示UAP56和COP1可以直接结合到剪接体和靶基因的mRNA上,表明UAP56和COP1可以通过作用于剪接体从而直接调控光信号介导的选择性剪接。
该研究工作一方面鉴定到光形态建成的新成员UAP56,另一方面发现了COP1作为可变剪接调节因子的新功能,揭示了UAP56与COP1共同通过调节下游基因可变剪接来调控光形态建成的分子机制和调控网络,增加了对植物光信号转导机制的认识。同时,UAP56在植物、酵母、动物等都广泛存在,具有保守的RNA解旋酶结构,UAP56-COP1的作用方式为研究其它物种的类似机制提供了重要参考。
该研究成果于2022年8月3日在线发表于国际学术期刊The Plant Cell上。植物所已毕业的李洋博士为第一作者,林荣呈研究员为通讯作者。该研究得到国家重点研发计划、国家自然科学基金以及中科院项目的支持。
文章链接:https://doi.org/10.1093/plcell/koac235
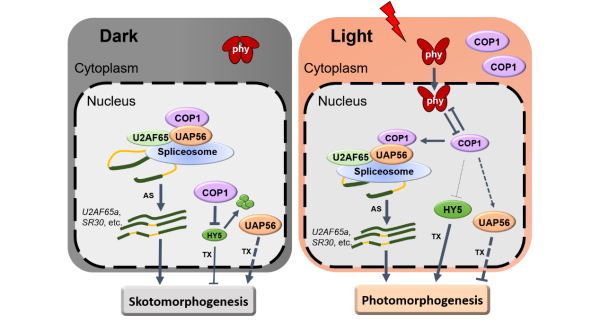
UAP56-COP1与剪接体互作调控RNA可变剪接进而影响光暗形态建成的工作模型
(光合实验室供稿)